Chlorophyll
Chlorophyll is the primary photosynthetic pigment class and its structure reveals the mechanism of its absorption of light, the excitation of its electrons, and its ability to give electrons off to an electron transfer system. Chlorophyll absorbs photons at wavelengths of between 400 and 700nm and channels their energy into the process of photosynthesis. The molecules of chlorophyll (C55H70MgN4O6) are large. Instead, they are attached to the membranes of disc-like structures, called chloroplasts, inside the cells.
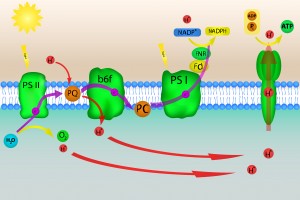
Photosynthesis occuring in chloroplast of plants
Chloroplasts are the site of photosynthesis, the process in which light energy is converted to chemical energy. The photons travelling along the waves enter the thylakoid space within chloroplasts. The light absorbed by chlorophyll space (which lines the inside of the thylakoid) supplies the energy used by plants to transform carbon dioxide and water into oxygen and carbohydrates. It will have a general formula of Cx(H2O)y. Basically, the chlorophyll vibrates because of the energy from the trapped photon. It can then transfer the photon to be used as energy.
| x CO2 + y H2O | light | x O2 + Cx(H2O)y |
| chlorophyll |
This is a chemical change that is accompanied by absorption of heat (endothermic transformation). The energy of the light absorbed by chlorophyll is converted into chemical energy stored in carbohydrates (sugars and starches). This chemical energy drives the biochemical reactions that cause plants to grow, flower, and produce seed.
There are several kinds of chlorophyll, the most important being chlorophyll a. This is the molecule which makes photosynthesis possible, by passing its energized electrons on to molecules which will manufacture sugars. All plants, algae, and cyanobacteria which photosynthesize contain chlorophyll A. Chlorophyll A(lpha) is sensitive to spectral peaks at 430nm and 660nm while chlorophyll B(eta) is sensitive to spectral peaks ranging around 460nm and 630nm. In order to absorb and transmute as much of the light’s given spectral energy as possible, pigments other than green 530nm are also required. But a few wavelengths between 500 and 575 nm are fairly effective for photosynthesis. The lower yield for the 500–575nm range of the spectrum explains the green color of plants. These wavelengths are reflected and transmitted rather than being as effective in driving the process. The absorption spectrum is for a chloroplast, antenna pigments are at work here. These ancillary pigments cannot directly convert sunlight into sugar and hence must pass their energy to the chlorophyll receptors for final conversion. Literally speaking, the plant essentially captures the light bands that chlorophyll cannot.
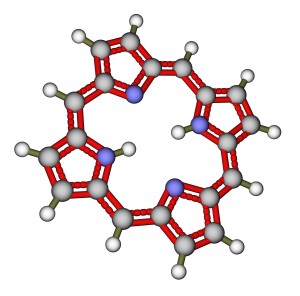
Porphyrin Ring
Chlorophylls are greenish pigments which contain a . Chlorophyll A is a type of porphyrin, which is the chemical that gives plants and leaves their green colour. This is a stable ring-shaped molecule around which electrons are free to migrate. Because the electrons move freely, the ring has the potential to gain or lose electrons easily, and thus the potential to provide energized electrons to other molecules. This is the fundamental process by which chlorophyll “captures” the energy or photons provided by the way of a grow light or the sun.
Plants require warm temperatures and sunlight to produce chlorophyll in autumn. The amount produced begins to decrease, and the existing chlorophyll is slowly broken down, diminishing the green colour of the leaves. Beta-carotene is the most abundant with a strong preference for blue light while other carotenoids absorb bands of yellow, orange and red.
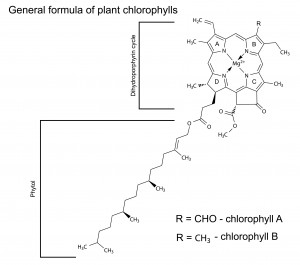
Formula of Plant Chlorophyll’s
Chlorophylls are the primary photosynthetic pigment class as they reflect green and absorb blue and red wavelengths. Chlorophyll A is the primary electron capture to drive the process of generating carbohydrates for long term storage of energy.
Accessory pigments can include chlorophyll B, xanthophylls, and carotenoids (such as beta-carotene). They absorb energy from spectra that chlorophyll A does not absorb and passes it to chlorophyll B.
Chlorophyll A with accessory pigment chlorophyll B are in light harvesting complexes in plants. The main function of chlorophyll is to collect energy; it’s a secondary function which is to regulate the light harvesting antenna size.
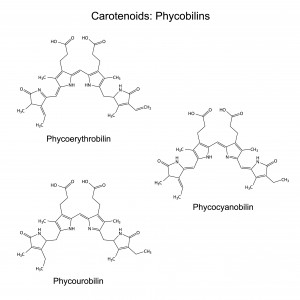
Carotenoids : Phycoblins
Carotenoids absorb light in the blue–green and violet region and reflect the longer yellow, red, and orange wavelengths. Carotenoids are composed of two small six-carbon rings connected by a chain of carbon atoms. As a result, they do not dissolve in water, and must be attached to membranes within the cell. Carotenoids cannot transfer sunlight energy directly to the photosynthetic pathway, but must pass their absorbed energy to chlorophyll. For this reason, they are called accessory pigments.
Karrer,(Karrer,1934; Karrer and Helfenstein, 1933) had recognized the symmetrical nature of the various carotenoids (b -carotene; lycopene; zeaxanthin) and that vitamin A was related to half of the b -carotene molecule.
Carotenoids are known to have another major function that of which is called ‘photoprotection’ of reaction centers, pigment-protein antennae, and cells and tissues (see Krinsky, 1968, 1979). Carotenoids absorb at approximately 480nm thus they are are universally associated with photosynthetic systems because they protect these systems against photodynamic damage.
There are over 600 known carotenoids; they are split into two classes, xanthophylls (which contain oxygen) and carotenes (which are purely hydrocarbons, and contain no oxygen). They serve two key roles in plants and algae: (1) they absorb light energy for use in photosynthesis, and (2) they protect chlorophyll from photo-damage. In other words, carotenoids collect light in wavelengths that chlorophyll can’t and then transfer it to chlorophyll. They also absorb excess light energy and dissipate it in order to avoid damage in what is termed the Xanthophyll Cycle. In addition to the xanthophyll cycle pigments (zeaxanthin and antheraxanthin), α-carotene-derived xanthophylls such as lutein, which are structural components of the subunits of the light-harvesting complexes, contribute to the dissipation of excess absorbed light energy and the protection of plants from photo-oxidative damage.
Carotenoids : Xanthophylls
Chlorophyll has some help in the form of the pigments carotene and xanthophyll, which in short, Xantho is Greek for ‘yellow’. Carotene (‘kar-uh-teen) is what gives organic items like egg yolks their orangish colour. These two pigments are always present in leaves and help absorb sunlight , which they transfer to chlorophyll for photosynthesis. In most plants, the yellow colouration is masked by the presence of chlorophyll, and xanthophylls are only revealed when chlorophyll is degraded when the leaf has become senescent. These yellow pigments are also observed in leaves with certain nutrient deficiencies such as chlorosis.
Carotenoids : Carotenes
B-Carotein, (C40H36) is a class of carenoids which contribute orange colours. Beta-carotene is one of the most common carotenoids in plants, and absorbs green and blue light strongly, reflecting red and yellow light and causing its orange appearance. It is also responsible for the orange colouration of carrots. Carotene, (C40H36) is a large molecule which is contained in the chloroplasts of many plants.
Pumpkins change color from green to orange for the same reasons tree leaves change color in the fall, and they do it under the same conditions. Most pumpkins contain organic pigments called carotenoids that give their flesh and skin the classic deep orange tint. Not all pumpkins turn orange, though some are selectively bred to be white, red or even blue.
Even some green vegetables, including spinach and broccoli, have large amounts of carotenoids hiding under all that deep green chlorophyll. Light energy absorbed by carotenoids is transferred to chlorophyll a and used in photosynthesis. In support of this it has been show that light that is absorbed by carotenoids 450nm results in chlorophyll flourescence 680nm. Carotenoids also play an important role in photoprotection (Demmig-Adems and Adems 1996). When chlorophyll is activated by the absorption of light it is generally returned to its original state as a result of its participation in photosynthesis. Absorption of access light can destroy chlorophyll in a process called photo-bleaching. These extremely reactive singlet oxygen species exist as free radicals(molecules with one or more unpaired electrons) which can oxidize or bleach organic compounds such as chlorophyll.
Carotenoids in leaves start degrading at the same time as chlorophyll, but they do so at a much slower rate; beta-carotene is amongst the most stable, as some fallen leaves can still contain measurable amounts.
Anthocyanins are unlike the carotenoids. Anthocyanin synthesis is kick-started by the onset of autumn – as sugar concentration in the leaves increases, sunlight initiates anthocyanin production. The purpose they serve isn’t clear, but it’s been suggested that they help protect the leaves from excess light, prolonging the amount of time before they fall.
One of these changes is the growth of a corky membrane between the branch and the leaf stem. This membrane interferes with the flow of nutrients into the leaf. As a result of the nutrient flow being interrupted, the production of chlorophyll in the leaf declines, and the green colour of the leaves fade.
When it forms this separation layer made of cork-like cells at the base of each leaf, it seals it off from the tree. It stops producing chlorophyll since it won’t need this pigment until the days start to lengthen once again in the spring. When the abundance of chlorophyll diminishes, the yellow and orange pigments get a chance to shine.
The red hues, which come from pigments called anthocyanins, are slightly more complicated. Whereas all trees contain chlorophyll, carotene and xanthophyll, not all of them produce anthocyanins. Even the ones that do have anthocyanins only produce it under certain circumstances. Two prevailing theories are that anthocyanins protect the leaves from excess sunlight and enable the trees to recover any last remaining nutrients. Secondly, another possible reason has been proposed: when the leaves decay, the anthocyanins seep into the ground and prevent other plant species from growing in the spring.
The bright red and purple fall foliage colours come from anthocyanin (an-thuh-‘si-uh-nuhn) pigments. These are potent antioxidants common in many plants; for example, beets, red apples, purple grapes (and red wine), and flowers like violets and hyacinths.
Anthocyanins
The layer of cells at the base of the leaf has one sole purpose which is to protect the tree during the colder winter and prevent it from drying out. When the separation layer is complete, the leaves fall off in the tree’s attempt to conserve energy. But before the leaves fall off it attempts to pull in as much sugar and nutrients as possible from its leaves, which is where the anthocyanin comes in.
Anthocyanins, are a accessory pigment class that reflects red, blue, purple, or magenta colours. Recent research shows that there are 550 known anthocyanins but these are not involved in photosynthesis. They give colour to flowers & fruit and are there for protection from UV or oxidative damage.
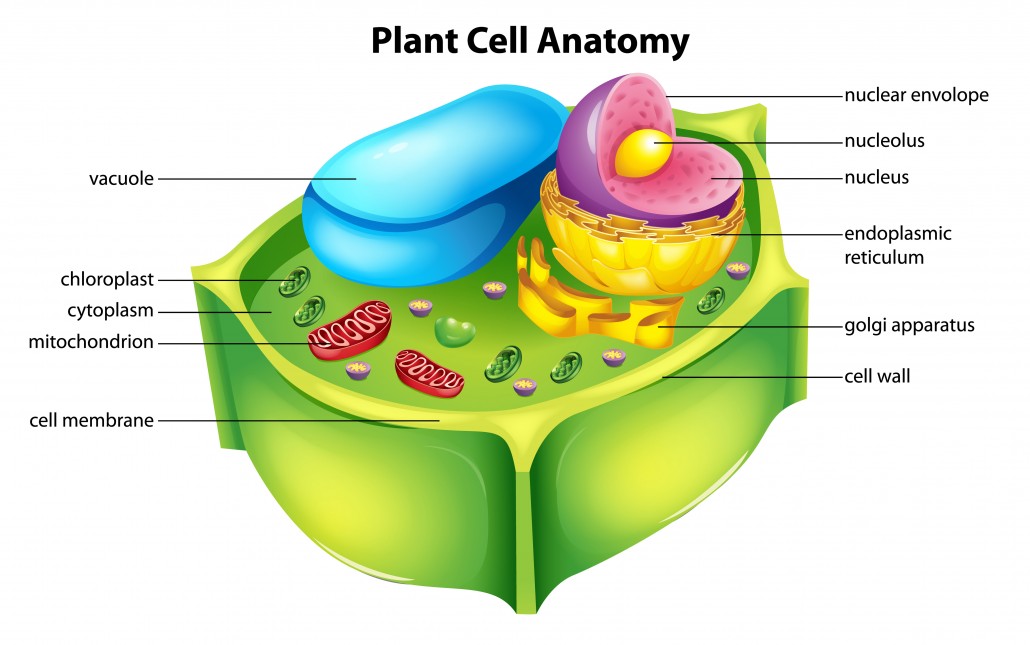
Plant cell Anatomy
Plants can sense and respond to different stimuli such as chemical concentrations in the air and soil, to water, from being touched, any motion or vibration that may be present in the environment around them, pathogens, predators and also light and spectra heavily influence their development.
“Recent biochemical and molecular work is producing a growing list of elements involved in responses to biotic and abiotic stimuli that are very similar across kingdoms. Some of the more interesting examples of these include prostaglandin/octadecanoid-mediated responses to wounding, steroid-based signaling systems, and pathogen-recognition mechanisms. Some of these similarities probably represent evolutionary convergence; others may be ancestral to plants and animals. Ecological and evolutionary implications of such overlaps include the existence of pathogens that can cause disease in plants and animals, the ability of herbivores to manipulate plant responses, usurpation of microbial mechanisms and genes by herbivorous animals and plants, evolution of plant defenses exploiting shared signals in animals, and the medicinal use of plants by humans. Comparative study of the signaling and response mechanisms used by plants, animals, and microbes provides novel and useful insights to the ecology and evolution of interactions across kingdoms.”
When working with LEDs to grow and cultivate the wide variety if plant life that we consume one begins to realize that there are vast variables which we are just beginning to understand when it comes to the relationship between plants and animals. It has been suggested that plants and animals share the same molecular pathways in order to respond to stress, it is convincing to say the least that a molecule produced in plants can also be effective in animals. Plants create a wide array of protective substances some of which can induce similarly protective responses in those who eat them.




